#eretmorhipis
Text
Being underwater can be relaxing, you get to have a moment of peace where all you see is an endless blue... until something blocks the view
I'm Hilda's case, however, it's a rather odd sight: an Eretmorhipis; this one it's on its way to the shoreline, perhaps to lay it's eggs, or simply to rest for the night
Who knows?


2 notes
·
View notes
Text

Hupehsuchians were small marine reptiles closely related to ichthyosaurs, known only from the Early Triassic of southwestern China about 249-247 million years ago. They had toothless snouts, streamlined bodies, paddle-like limbs, and long flattened tails, along with a unique pattern of armor along their backs made up of overlapping layers of bony osteoderms.
Hupehsuchus nanchangensis was a mid-sized member of the group, about 1m long (3'3"). Newly-discovered fossils of its skull show that its long flattened snout had a distinctive gap between the bones (similar to the platypus-like snout seen in its relative Eretmorhipis) with an overall shape surprisingly convergent with that of modern baleen whales – suggesting that this hupehsuchian may have been a similar sort of filter-feeder.
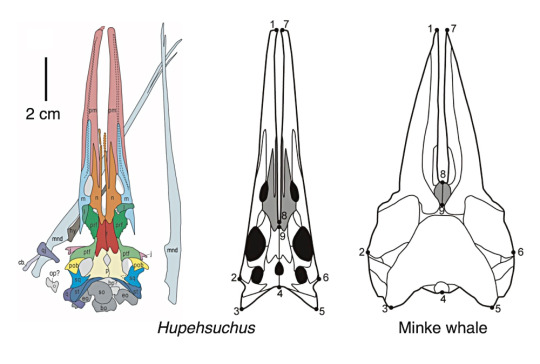
Hupehsuchus skull compared to a modern minke whale
[From fig 2 & fig 3 of Fang et al (2023). First filter feeding in the Early Triassic: cranial morphological convergence between Hupehsuchus and baleen whales. BMC Ecol Evo 23, 36. https://doi.org/10.1186/s12862-023-02143-9]
Grooves in the bones along the outer edges of its upper jaws may be evidence of filtering structures similar to baleen, although with no soft-tissue preservation we don't know exactly what this would have looked like. Its slender flexible lower jaws probably also supported a large expandable throat pouch, allowing it to filter plankton out of larger volumes of water.
———
NixIllustration.com | Tumblr | Patreon
#science illustration#paleontology#paleoart#palaeoblr#hupehsuchus#hupehsuchia#ichthyosauromorpha#reptile#marine reptile#art#triassic weirdos
596 notes
·
View notes
Text
Eretmorhipis carrolldongi

Art by: Liam Elward, https://www.deviantart.com/prehistorybyliam/art/Eretmorhipis-783558933
Name: Eretmorhipis carrolldongi
Name Meaning: Oar fan
First Described: 2015
Described By: Chen et al.,
Classification: Chordata, Tetrapoda, Reptilia, Hupehsuchia, Hupehsuchidae
Eretmorhipis was a weird and very strange marine reptile. It had osteoderms on its back, thick ribs, and its digits were fan shaped. Eretmorhipis is actually a part of Hupehsuchia, which are pretty much reptiles that are close in relation to ichthyosaurs. Anyways, Eretmorhipis existed during the Early Triassic and was found in China’s Jialingjiang Formation. We’ve found specimens in the past that consisted of post-cranial material, but most recently in 2018, paleontologists found a specimen that included the skull and mostly complete post-cranial remains of this odd aquatic creature.
Sources:
https://en.wikipedia.org/wiki/Eretmorhipis
#Eretmorhipis#palaeoblr#paleontology#palaeontology#prehistoric#ancient marine reptiles#Mesozoic#Not a dinosaur#ichthyopterygia
28 notes
·
View notes
Link
2 notes
·
View notes
Text
Hupehsuchus nanchangensis

By Scott Reid
Etymology: Hubei crocodile
First Described By: Young & Dong, 1972
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Disapsida, Neodiapsida, Ichthyosauromorpha, Hupehsuchia, Hupehsuchidae
Referred Species: H. nanchangensis
Status: Extinct
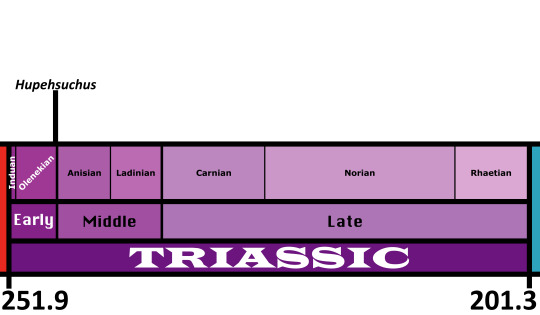
Time and Place: Between 248 to 247 million years ago, in the Olenekian of the Early Triassic
Hupehsuchus is known only from the Jialingjiang Formation in the Hubei Province of China.
Physical Description: At first glance, Hupehsuchus looks a lot like what you’d expect a primitive ichthyosaur to look like, and indeed they are related. Only around 1-2 metres (3-6 feet) long, its body is long and streamlined, with a long, pointed snout lacking teeth, limbs evolved into large rounded paddles, and a long flattened tail without a fluke. However, on closer inspection Hupehsuchus turns out to be much more bizarre.
Like other hupehsuchians, Hupehsuchus is heavily armoured with vertically layered rows of osteoderms running down its back over the vertebrae (you read that right, they’re on top of each other). These osteoderms interlock with each other, which may have stiffened the body and/or provided ballast. The vertebrae themselves have tall neural spines, giving Hupehsuchus an almost humpbacked appearance, and the neural spines appear to be made up of two separate pieces, one attached to the vertebra, the other seemingly attached to the osteoderms. Only hupehsuchians have vertebrae like that. Its body was probably compressed from side to side, taller than it was wide, and was only flexible around the hips and tail. The armour on its back is complemented by a robust, thick ribs that almost create a shield around its sides, and it even has heavily built, interlocking gastralia on its underside complete with another row of smaller osteoderms on the underside.
The skull of Hupehsuchus is also very unusual. The jaws are long, pointed, and the skull was seemingly quite broad and flat from above, resembling a duck’s bill. The lower jaws were incredibly thin and loosely attached to each other, and were probably capable of bending and bowing outwards like a pelican or baleen whale. The lower jaw may even have supported a large gular pouch, and large hyoid bones imply it had a powerful tongue. Furthermore, several parallel grooves in the upper jaw suggest the presence of a baleen-like structure in its mouth, making it even more uncannily similar to baleen whales. The neck, though, is relatively long and slender compared to a whale.
Diet: The bizarre adaptations of Hupehsuchus suggest it may have been a filter-feeder, sifting small particles and animals out of the water column or near the sea bed.
Behavior: Hupehsuchus was likely a lunge-feeder, in spite of its relatively long neck. The laterally compressed body and flexible hips and tail indicate that it swam by undulating through the water, likely lunging forwards suddenly through swarms of plankton and other small organisms to feed. The flexible jaws of Hupehsuchus imply that it engulfed large amounts of water as it fed, like whales, and would have strained it out through its filtering structures using its tongue.
Hupehsuchus almost certainly gave birth to live young, like ichthyosaurs. Partly because its anatomy was totally unsuited for crawling onto land to lay eggs, but also because one of the oldest ichthyosaurs, Chaohusaurus, is known to have given birth to its babies head-first, the opposite of the tail-first birth in other ichthyosaurs. Babies born head-first in water are at risk of drowning during birth, so this implies that live birth evolved in the ancestors of ichthyosaurs while they were still on land, which would mean that Hupehsuchus likely would have inherited this trait too.
Ecosystem: Hupehsuchus, and indeed all other hupehsuchians for that matter, are known only from a single locality in China. Here Hupehsuchus coexisted with its fellow hupehsuchians Nanchangosaurus, little short-necked Eohupehsuchus, the tubular Parahupehsuchus (I’m sensing a pattern here) and the truly bizarre platypus-faced Eretmorhipis (along with an undescribed polydactylous species!).
The diversity of hupehsuchians in this one habitat likely lead to diverse niche partitioning between them. Hupehsuchus was one of the largest, and seemingly occupied a more active lifestyle, lunge-feeding on organisms in the water. Nanchangosaurus had a similar skull to Hupehsuchus, and so may have had a similar diet, but it was smaller and had a body shape more suited for swimming along the sea bed. Parahupehsuchus was so heavily encased in armour that its body was practically a bony tube, and rather than round paddles it had pointed flippers, suggesting a different swimming style (for what, we don’t know). The bizarrest of them all, Eretmorhipis, may have specialised in grubbing blindly through the seabed, sensing prey with its bill like a platypus, possibly even at night.
The habitat is believed to have been a shallow lagoonal environment, perhaps sheltered from larger predators that could have preyed upon them, like giant nothosaurs. However, they nonetheless coexisted with the primitive ichthyosaur Chaohusaurus and two species of pachypleurosaur (sauropterygians related to nothosaurids), Keichousaurus and Hanosaurus, as well as an undescribed large sauropterygian 3-4 metres (10-13 feet) long. Strangely, no fish have been discovered in this formation, so it’s quite possible that the smaller hupehsuchians were the main food source for some of these larger marine reptiles (as evidenced by a bite taken out of one Eohupehsuchus paddle!). These conditions may have also prompted the strange dietary adaptations of the hupehsuchians, with no smaller fishes to eat, they specialised in eating tiny marine invertebrates and other plankton.
Other: Hupehsuchians are very mysterious marine reptiles, they are known from only one location in the whole world and from a very narrow range of time in the earliest Triassic. Hupehsuchus was once suggested to be a missing link between the ichthyosaurs and their as-yet-unknown terrestrial ancestors, although as more early ichthyosauromorphs have been discovered it is clear that is not the case, and that hupehsuchians are a bizarre offshoot of their own.
Hupehsuchus is part of a surprisingly diverse range of early-derived ichthyosauromorphs that lived in China during the Early Triassic, just a few million years after the Permian Mass Extinction, including the first proper ichthyopterygians and the peculiar (possibly amphibious) nasorostrans like Cartorhynchus. These marine reptiles were very quick to diversify in the wake after the extinction, as the strange filter-feeding lifestyle of Hupehsuchus testifies, quite the opposite of the predicted slow recovery for marine ecosystems. However, it remains a mystery why only the ichthyosaurs prevailed, and all the other strange and diverse ichthyosauromorphs like Hupehsuchus never even made it into the Middle Triassic. Perhaps they were just too strange and specialised even for the Triassic.
~ By Scott Reid
Sources under the Cut
Carrol, Robert L.; Dong, Z.-M. (1991). "Hupehsuchus, an enigmatic aquatic reptile from the Triassic of China, and the problem of establishing relationships". Philosophical Transactions: Biological Sciences. 331 (1260): 131–153.
Chen, X. H.; Motani, R.; Cheng, L.; Jiang, D. Y.; Rieppel, O. (2014). "A Carapace-Like Bony 'Body Tube' in an Early Triassic Marine Reptile and the Onset of Marine Tetrapod Predation". PLoS ONE. 9 (4): e94396.
Chen X. H., Motani, R., Cheng, L., Jiang, D. Y., Rieppel, O. (2014) “The Enigmatic Marine Reptile Nanchangosaurus from the Lower Triassic of Hubei, China and the Phylogenetic Affinities of Hupehsuchia”. PLoS ONE 9(7): e102361.
Chen, Xiao-hong; Motani, Ryosuke; Cheng, Long; Jiang, Da-yong; Rieppel, Olivier (May 27, 2015). "A New Specimen of Carroll's Mystery Hupehsuchian from the Lower Triassic of China". PLoS ONE. 10 (5): e0126024.
Cheng, L., Motani, R., Jiang, D.Y., Yan, C.B., Tintori, A. and Rieppel, O., (2019). “Early Triassic marine reptile representing the oldest record of unusually small eyes in reptiles indicating non-visual prey detection”. Scientific reports, 9(1), p.152.
Motani, R., Chen, X. H., Jiang, D. Y., Cheng, L., Tintori, A., Rieppel, O. (2015). “Lunge feeding in early marine reptiles and fast evolution of marine tetrapod feeding guilds”. Scientific reports. 5: 8900.
Wu, X.-C.; Li, Z.; Zhou, B.-C.; Dong, Z.-M. (2003). "A polydactylous amniote from the Triassic period". Nature. 426 (6966): 516.
Xiao-hong Chen; Ryosuke Motani; Long Cheng; Da-yong Jiang & Olivier Rieppel (2014). "A Small Short-Necked Hupehsuchian from the Lower Triassic of Hubei Province, China". PLoS ONE. 9 (12): e115244.
Xiao-hong Chen; Ryosuke Motani; Long Cheng; Da-yong Jiang; Olivier Rieppel (May 27, 2015). "A New Specimen of Carroll's Mystery Hupehsuchian from the Lower Triassic of China". PLoS ONE. 10 (5): e0126024.
#Hupehsuchus nanchangensis#Hupehsuchus#Triassic#Ichthyosaur#Reptile#Marine Reptile#Prehistoric Life#Palaeoblr#Paleontology#Prehistory#Triassic Madness#Triassic March Madness
551 notes
·
View notes
Link


An article published in the journal "Scientific Reports" reports the discovery of two new specimens of Eretmorhipis carrolldongi, a marine reptile that lived about 250 million years ago, in the Early Triassic period, in today's China. A team of researchers studied these new specimens with particular interest because of the previous ones one had its skull but it was in a poor state of conservation and the other one had no skull. The new specimens show that this reptile had similarities with modern platypuses, with a sort of bill of cartilage and receptors that allowed it to hunt based on touch.
0 notes
Note
Eretmorhipis was a strange aquatic reptile that lived about 250 million years ago. It had a very stiff body and fail, unusually for an aquatic animal, and possessed crocodile like back armor. It’s strangest feature was its head, which was platypus like, indicating it may have been doing platypus like things (ie searching for food by touch and electroreception in murky water)

Ooooooooh, interesting :ooo
2 notes
·
View notes
Photo

A fossil of Eretmorhipis carrolldongi, a marine reptile with a tiny head and large paddle-like limbs.
Like the platypus, this recently discovered prehistoric creature had a duckbill. But then nature made it even weirder, adding plates on its back like a stegosaurus, a long tail like a crocodile, large paddle-like limbs and a tiny head with teeny eyes. This specimen dates to about 247 million years ago during the early Triassic, a time before dinosaurs ruled Earth.
Credit: Long Cheng and Ryosuke Motani
63 notes
·
View notes
Link
My, what tiny eyes you had, Eretmorhipis carrolldongi.
Two newly found specimens of the mysterious, platypus-like reptile suggest that the ancient creature had very small eyes for its size, and may have hunted mainly by touch. That makes E. carrolldongi the oldest known amniote — a group that includes reptiles and mammals — to use a sense other than sight to find its prey, scientists report online January 24 in Scientific Reports.
E. carrolldongi, which lived about 250 million years ago, is one of numerous strange creatures dating to the Early Triassic described by scientists in recent years. It is part of an oddball array of marine reptiles called Hupehsuchia that lived in a vast lagoon spanning hundreds of kilometers across what’s now southern China. That flourishing of forms, which came on the heels of the mass extinction at the end of the Permian Period 252 million years ago, suggests that marine reptiles diversified millions of years earlier than once thought, the researchers say.
E. carrolldongi was named in part for its large, fan-shaped flippers, which give its body a passing resemblance to a platypus (Eretmorhipis means “oar fan”). Now, the newly discovered specimens, the first with skulls, point to one more thing that the ancient swimmer had in common with the modern platypus: very small eyes.
The creature also had a small head, meaning that it probably didn’t use hearing to forage, given the challenge of localizing sound in water. Chemoreception — used by snakes, for example, to gather information from the atmosphere through their tongues — is also unlikely based on the lack of certain telltale holes the skull, say paleontologist Long Cheng of the Wuhan Centre of China Geological Survey and colleagues.
By elimination, the researchers suggest that E. carrolldongi probably used tactile cues, such as hair cells that can help an animal detect movement, to stalk its lagoon prey. Still, electroreception, in which predators sense electric fields generated by moving prey, can’t be ruled out, the scientists say. And that would be one more thing it had in common with platypuses — they use electroreception, too.
4 notes
·
View notes
Photo


Cina, scoperto fossile di un rettile del Triassico simile ad un ornitorinco
La struttura di Eretmorhipis carrolldongi ricorda quella del mammifero semi-acquatico e probabilmente si procurava il cibo allo stesso modo, frugando i fondali con il becco
di Eleonora Degano
2 notes
·
View notes
Text
My favorite part
Ichthyosauromorpha (Motani et al., 2015a) is an expansive group that includes the two clades Hupehsuchia (Young & Dong, 1972b) and Ichthyosauriformes. Hupehsuchia is a distinctive clade of Early Triassic marine reptiles so far known only from five southern Chinese species, namely Nanchangosaurus suni (Wang, 1959), Hupehsuchus nanchangensis (Carroll & Dong, 1991), Parahupehsuchus longus (Chen et al., 2014b), Eohupehsuchus brevicollis (Chen et al., 2014c) and Eretmorhipis carrolldongi (Chen et al., 2015).
0 notes
Photo

Eretmorhipis carrolldongi, a hupehsuchian marine reptile from the Early Triassic of China (~247 mya).
This species was originally named back in 2015, but at the time the only known specimens were missing their heads. It was assumed that its skull would have looked similar to those of other hupehsuchians... but now new fossils have been found, and it seems to have actually been much much weirder!
Eretmorhipis’ head was surprisingly tiny in proportion to its body -- sort of like a marine version of Cotylorhynchus -- and its shape convergently resembled the modern platypus, with a wide “duck bill” and very small eyes. It may have hunted for food along the seafloor in a similar manner to the platypus, using either a highly sensitive sense of touch or possibly even electroreception to locate small invertebrates like worms and shrimp.
It also had much larger bony osteoderms than its other known hupehsuchian relatives, forming a distinctive protruding spiky ridge down its back. At about 85cm in length (2′9″) it was one of the largest marine animals around at the time, so this structure probably wasn’t needed for defense -- but as with other hupehsuchians its actual function is still unknown.
#science illustration#paleontology#paleoart#palaeoblr#eretmorhipis#hupehsuchia#ichthyosauromorpha#marine reptiles#triassic#it's a hupehsuchian of course it's weird#art#duck bills (woo ooh!)#i love hupehsuchians so much and i hope they keep on surprising us
422 notes
·
View notes
Text
Hupehsuchus nanchangensis

Art by: Smokeybjb, CC BY-SA 3.0
Name: Hupehsuchus nanchangensis
Name Meaning: Hubei crocodile (Hubei Province in China)
First Described: 1972
Described By: Young and Dong
Classification: Chordata, Tetrapoda, Reptilia, Hupehsuchia, Hupehsuchidae
Remember my post on Eretmorhipis, the weird and strange guy from the Early Triassic who was a part of a clade of marine reptiles that were closely related to ichthyosaurs? Well, we have another member of that clade for today’s entry! Hupehsuchus was also from the Early Triassic and was discovered in China and they too were pretty strange in their own right. They had a long toothless snout, long tail, and their limbs were like paddles, but not fully formed flippers as seen in later ichthyosaurs. Oh and let’s not forget, Hupehsuchus had armor on its back as well. There’s still a debate within the paleontological community on whether Hupehsuchus was even closely related to the ichthyosaurs or not. Some hypothesize that because this guy had extra bones within its hands instead of within its individual fingers, that the difference itself may possibly rule Hupehsuchus out in close relation to ichthyosaurs.
https://en.wikipedia.org/wiki/Hupehsuchia
https://en.wikipedia.org/wiki/Hupehsuchus
#Hupehsuchus#palaeoblr#paleontology#palaeontology#prehistoric#Mesozoic#ancient marine reptiles#Not a dinosaur#ichthyopterygia
27 notes
·
View notes
Photo

This is Eretmorhipis carrolldongi, the reptilian platypus from the Early Triassic of China... https://www.instagram.com/p/CZqEtNmJzvd/?utm_medium=tumblr
1 note
·
View note
Text
Episode 5: A World of Ruling Reptiles

Image credit: Mark Witton, under CC BY 2.0.
The following is the transcript for the fifth episode of On the River of History.
For a link to the actual podcast, go here. (Beginning with Part 1)
Part 1
Greetings everyone and welcome to episode 5 of On the River of History. I’m your host, Joan Turmelle, historian in residence.
The survivors of the great mass extinction event at the end of the Permian inherited an impoverished world. With up to 96% of all organisms having kicked the bucket, there were essentially no ecosystems present. The land was scorched and charred by a hot-house climate and the seas chocked with acidity. It took over 10 million years for life to recover, and when it did, the planet again hosted a pageant of strange and fantastic creatures, shuffled along by whatever environmental changes came their way.
This episode concerns the Mesozoic Era, which began with the close of the Paleozoic 251.9 million years ago, and lasted until 66 million years ago. The name Mesozoic means ‘middle life’, which is fitting considering it’s sandwiched between the other two eras of the Phanerozoic Eon.
The Earth at the beginning of the first period of the Mesozoic, the Triassic (251.9 million to 201.3 million years ago), was still dominated by the supercontinent of Pangaea. Nearly all the landmasses of the Earth were connected together, but as the Earth’s tectonic plates are constantly moving at a rate of centimeters per year, there were changes on the horizon. For one, the great Paleo-Tethys ocean was slowly closing as the eastern most fringe of Pangaea called Cimmeria (land that included present-day East and Southeast Asia) was moving upwards towards the greater landmass. This was taking the Paleo-Tethys with it, closing it up and making room for a new ocean to form, the Tethys proper. Throughout the Triassic, Pangaea as a whole was moving northwards and this placed the newborn Tethys ocean along the equator. This meant that the seasons became more pronounced and violent and the years were punctuated by periods of harsh wet rains and vast dry periods.
The marine environments of the world started the Triassic as diminished communities of mollusks and crustaceans, meekly trailing the seabed in search of food. They were much smaller than their predecessors: a consequence of mass extinctions. For this brief period of time, stromatolites made a comeback as the hordes of grazing invertebrates that previously eliminated their number were themselves weakened and no longer a major threat. But the recovery of species soon found this collection of marine organisms in a much better state. Reef-building organisms returned and on the evolutionary stage sprang a new group of corals, the scleractinians. Though the name may be new to you, these animals certainly aren’t, because many modern corals alive today belong to this lineage. Like their now-extinct rugose and tabulate relatives, scleractinian corals (or stony corals) can form colonies or remain singular organisms, but unlike them their skeletons are much softer and more permeable. Early coral reefs started out very small, and it wouldn’t be until later in the Triassic that they gained more prominence.
The coiled ammonoids fully recovered following the Permian extinction event and very quickly diversified into hundreds of new species. Bivalve and gastropod mollusks increased in numbers as well and continued to push away their brachiopod neighbors, who themselves remained small and unimportant members of the oceans for the remainder of the Phanerozoic. Among deuterostomes, echinoderms were doing well, with the filter-feeding crinoids and hard-plated urchins experimenting with new forms. The fishes had bounced back as well: in particular the sharks and ray-finned fishes. Lobe-finned fishes never recovered after the Permian, and only two lineages survived on to the present day. These were the lungfishes, who are freshwater organisms specialized for oxygen-poor waters, and the coelacanths, who are marine fishes that originally came in a variety of body forms.
Moving along to near-shore waters and terrestrial habitats, reptiles were experiencing a boost in biodiversity. At the start of the Triassic, the Earth was originally in the grasp of the therapsid protomammals, of which a few groups had survived the extinction. There were the pig-like, tusked dicynodonts that feasted on woody plants; there were the carnivorous therocephalians that preyed upon their larger relatives; and there were the cynodonts who may or may not have already sported a covering of fur and whiskers. But their reign wasn’t to last, as the reptiles that survived started to explore the new niches available to them and very gradually pushed the therapsids into obscurity. One area that reptiles excelled at was recolonizing the water. As previously mentioned, there already were semi-aquatic reptiles during the Permian Period, but it was in the Triassic Period that we started to see more of a push towards fully aquatic living. Some groups of nearshore reptiles include the placodonts, who adapted their teeth for crushing the tough shells of marine mollusks. This reliance on slow bottom-dwelling prey facilitated the development of heavy armor plating along their backs, which would have aided in their ability to stay on the seafloor for longer periods of time. Other reptiles underwent even stranger experiments, including Atopodentatus which sported a face like a vacuum-cleaner for digging through seafloor sands, Eretmorhipis which was essentially a reptile version of a platypus, and Tanystropheus which increased the length of its neck vertebrae so much that the animal was nearly all neck. The purpose of this exaggerated feature is currently debated, but a common interpretation sees the animal squatted along the seashore and using its neck like a fishing-pole, though free-swimming isn’t out of the question.
More typical among these new aquatic reptiles were the nothosaurs who were almost certainly capable of swimming in the open ocean after prey. Their heads sported thin teeth that would have made snagging slippery fish or squid a breeze. Descended from relatives of these reptiles were some of the first truly marine reptiles, the plesiosaurs which often sported long necks for catching fish. The keys to their success stem from two new traits. First, their limbs were flattened and completely encased in skin which formed paddles, like that on a canoe: this would have increased their efficiency in swimming. Second, plesiosaur ancestors abandoned egg-laying on land for live birth in the water. Newborns would have emerged tail-first and had to cruise quickly to the surface for their first breath of air. By changing the shapes of their limbs and becoming viviparous, plesiosaurs became dynamic marine reptiles. A possibly related lineage of reptiles took these adaptations to an even greater extreme. Ichthyosaurs, like plesiosaurs, evolved from amphibious, near-shore reptiles, but when they began to settle in the oceans they not only turned their limbs into fins and gained the ability of live-birth: they had streamlined their bodies into a fish-like shape (this, incidentally, is where they got their name). Now, alongside the sharks, the oceans had apex predators once again.
On land, the situation became just as pronounced. Like the earlier Permian, reptiles took on arboreal, burrowing, and gliding niches, but now they really started to acquire more important roles in their ecosystems. Among the lineages that survive to the present day there were three: the turtles which by the Late Triassic had managed to create their shells by solidifying their bodies in plates and moving their shoulders and limbs inside their ribcage; the lepidosaurs of which their descendant branches the lizards and tuataras had yet to diverge; and the archosaurs. It is this latter group that the Triassic blessed with unparalleled diversity.
The ancestors of the archosaurs were originally sprawling reptiles that bore resemblance to the lizards of today, but at some point in their lineage members took a page from the synapsids and switched to a more upright posture. This was supplemented by the addition of a hole in the skull between the eye socket and the nostril which appears to have housed an air-sac that would have lightened the skull as well as better regulate body temperature. It was here that two lineages diverged from each other around 247 million years ago: the pseudosuchians who mostly walked on semi-erect limbs and the ornithodirans who bore fully erect limbs underneath their bodies. These probably would have been endothermic organisms, able to generate their own body heat.
By the later part of the Triassic, these two groups of archosaurs were in competition with each other and it was the pseudosuchians who proved the more successful. They occupied nearly all of the major herbivorous and carnivorous niches in the later Triassic ecosystems. One group, the aetosaurs, bore bony armor along their backs and used their pointed snouts for eating low-growing plants. Another herbivore, Lotosaurus, was more of a pot-bellied animal that sported a beak (but no teeth) for nipping leaves. Among the carnivorous lot were the rauisuchoids who reached impressive sizes of 20 feet or more. Their large heads sported stabbing teeth and the structure of their limbs suggests that some may have been able to walk on their hind limbs for extended periods of time, possibly running down larger prey. A related group, the shuvosaurs, were obligate bipeds, walking on their hindlimbs all of the time.
With such large and intimidating reptiles roaming the land, the ornithodirans remained small-sized members of their communities, but they were experimenting too. By 228 million years ago, one lineage had achieved a major milestone in the history of vertebrates. These ornithodirans had developed flaps of skin between their bodies, supported by a greatly elongated finger (the equivalent to our ring finger). They became very light-weight and lengthened their heads while shortening their hindlimbs. These were the pterosaurs, that great group of reptiles that were the first vertebrates to develop true powered flight rather than gliding like earlier reptiles had done. By becoming airborne, pterosaurs were effectively removed from the competition going on below them, while the other ornithodirans continued to be pursued by the larger pseudosuchians. One group of these soon shifted their mode of locomotion towards proper bipedality, becoming more upright and lighter on their feet. Their digits changed, with the fingers being able to finally grasp prey items and the toes supporting the animal’s weight, rather than their soles. A notable change occurred in the hip bone, where the socket that the limbs attached to opened up completely. This allowed the animals to fully place their limbs underneath their bodies, which made them very efficient walkers and runners. It was roughly 233 million years ago that the dinosaurs entered the world.
There are perhaps no other prehistoric animals that have a greater pop-culture presence than the dinosaurs. Young children and elderly alike adore them, and they have inspired generations of people with their diversity and antiquity. Granted, as you have seen from our river journey, there were plenty of equally strange and often far more fascinating animals that have come and gone, but no matter what it is the dinosaurs that have stayed with us the most. So it might be surprising to know that dinosaurs were not properly described by science until the mid-1800s (just under 180 years ago). Their name is Greek and stems from two words meaning “fearfully-great reptiles”. This might give an impression that the researchers who first studied these animals were amazed by their teeth, claws, and horns, but this was rather a declaration of awe at their grandeur.
The first dinosaurs started out as small reptiles that appear to have been opportunistic omnivores. Over the remainder of the Late Triassic they had diversified into three lineages that each had their own distinctive anatomy and behavior. There were the theropods, of which the majority remained bipeds throughout their entire history. Their skulls were light-weight and their feet sported three prominent toes while the others were reduced or lost altogether. In their early days, theropods were predatory animals, mostly small in size but capable of reaching off-putting lengths: Liliensternus could grow to almost 17 feet long. The second group, the sauropodomorphs originally started out as very similar to the theropods (perhaps even in diet), but they were being separated by an increasing dependence on vegetation. Their digestive systems were modified to take a primarily plant-based diet, and their necks grew in length to allow for ease at food collection. This became reflected in their teeth as well, with the sharp, pointed shape being phased out for a more peg-shaped form. It wasn’t until the very tail end of the Triassic that the sauropodomorphs reached their characteristic grand sizes and a subgroup, the sauropods, dropped on all fours as their feet became columnar to support their weight. The biggest sauropods in the Triassic, like Isanosaurus and Antetonitrus, were around 20-30 feet long from their long necks to their long tails. The last group, the ornithischians, also developed into mostly herbivorous dinosaurs. On the tip of their lower jaw evolved the predentary bone, which formed a sort of beak for cutting plants, and their pubis bone (the lowest part of the pelvis) pointed backwards to allow more room for larger guts. Though this group must have evolved during the Triassic, their presence in the fossil record is lacking.
While these ruling reptiles were certainly important organisms, the land was also experiencing new evolution of land plants and animals. The gymnosperms and ferns of the Permian managed to bounce back and were the dominant land floras of the Triassic. New groups of conifers, cycads, and ginkgoes evolved across Pangaea, encouraged by periods of growth thanks to the intense wet seasons. A particularly beloved site in Arizona, USA is full of petrified wood from a family of trees called araucarias that evolved during the Triassic. These were massive trees, with some of the trunks sprouting up to 200 feet tall and sporting strange patterned leaves. In the undergrowth could be found the first frogs, who were not leaping amphibians but rather crawlers that still sported short tails. Early spiders developed spinnerets, which give species today the ability to produce silk into strands for webbing. Among the insects, the earliest two-winged flies or dipterans evolved (species in this group today may be familiar by the houseflies, horseflies, gnats, and mosquitoes). Also appearing were the sawflies, which sported a knife-like ovipositor on their abdomen for cutting into leaves, allowing them to lay their eggs. We’ll return to this herbivorous group of insects later on. Among the therapsids, one lineage of cynodonts became very reduced in size and began to shift to a more nocturnal existence as many of the minor reptiles began to feast upon them. These small, furry animals were the earliest mammals, born in the world as a seemingly insignificant part of the fauna. But they were tough survivors, and their switch to night-time living certainly did them a favor.
In an almost sadistic turn of events, the Triassic ended with another mass extinction event. As far as causation is concerned, this event has been tied to the beginning of rifting events in Pangaea as the supercontinent began to finally split apart. Fault lines started opening between the landmasses and there were volcanic eruptions associated with this. Conditions proved to be very poor for some of the organisms as carbon dioxide levels spiked. In the oceans there were the usual losses among marine invertebrates, with ammonoids doing particularly badly. Save for a few species like the early mammals, the therapsids met their end, and the great range of reptiles on land and near the seashore also died out. There would be no more aetosaurs, nothosaurs, rauisuchoids, placodonts, and other weird reptiles. That grand competition between pseudosuchians and ornithodirans was finally over, with the hands being dealt in the ornithodiran’s favor (even if they won by the process of elimination, literally). With the close of the Triassic came the assurance that the Mesozoic Era would be known as the age of dinosaurs.
Part 2
The next period of the Mesozoic was the Jurassic Period, 201.3 million to 145 million years ago. The rifting that seems to have triggered the Triassic mass extinction event continued at a slow pace. The ancestors of the modern continents were beginning to break apart for the first time, with the most dynamic change occurring along the center of Pangaea. Starting around 180 million years ago, deep rivers began to cut through the north of the supercontinent and split it into two major chunks: the long-lived Gondwana in the southern hemisphere and the new continent of Laurasia (composed of North America and Eurasia) in the northern hemisphere. The rivers widened more and more and became a great sea. This was the formation of the Atlantic Ocean, and towards the end of the period (around 148 million years ago) it was as wide as Saudi Arabia or western Europe. Speaking of Europe, the lands that would become this subcontinent had flooded and turned into a series of islands or an archipelago. It wouldn’t be until the end of the Jurassic that Gondwana itself started to rift apart, with the lands of Indomadagascar (India & Madagascar) and Sahul (Australia & New Guinea) moving southward as Africa and South America slowly began to split as well. The breakup of Pangaea marks the end of the great Panthalassic Ocean, now replaced by the Pacific Ocean. All the landmasses of the Earth were beginning to fragment into separate continents and this would dramatically affect the variety of living things. With the mostly homogenous faunas and floras now being separated, not just by deserts or mountains but by entire oceans, distinct biogeography was underway.
In the oceans, these changes were spearheaded by the evolution of new marine organisms. Most prominent was the replacement of key ocean phytoplankton. Phytoplankton – that is, photosynthesizing microbes that float in the water – were previously dominated by green algae. These relatives of land plants had suffered a heavy blow during the Permian and Triassic extinction events, and now that the general chemistry of the oceans had changed, and the levels of nutrients had shifted, their rule was being overthrown by the red algae who could better inhabit these new seas. Among these new phytoplankton were the dinoflagellates and coccolithophores. The former had certainly evolved early in the Proterozoic, but the later were new comers. Coccolithophores cover their bodies in thick plates called coccoliths (where they get their name), and when these plankton died their shells sank onto the seafloor and gradually became converted into a new form of limestone called chalk. Dinoflagellates are often well known today because some species form great red blooms in the seas that poison whatever mollusks happen to be present, which in turn can harm people who eat them.
Marine invertebrates were flourishing thanks to the change in plankton, which promoted the establishment of new ocean food webs. New groups of scleractinian corals and sponges formed small reefs along the warm shallow seas and provided homes for different animals, including the first tube worms, who form thin tubes made of minerals that protect their soft bodies while they filter-feed. Among the mollusks, the ammonoids increased in number and many forms grew to impressive sizes of 2 feet in diameter or more. It was in the Jurassic that the cephalopods who abandoned their outer-shells in place of interior shells (the belemnoids) or just lost them altogether (the octopodes and squids) became more important. Belemnoids in particular would have formed great swarms as they patrolled the seas in search of fish to eat, snagging them with mineralized hooks on their arms. From the shrimp-like decapods emerged the first true crabs and lobsters. Echinoderms continued to do well, and crinoids still formed great forests in some parts of the seas, though they were being overshadowed by the more common sea stars and urchins.
Marine vertebrates saw new developments too. By the middle of the Jurassic, around 170 million years ago, most of the major ray-finned fish lineages had evolved, including the ancestors of sturgeons, gars, herring, tarpon, eels, and carp. Among these fishes emerged a lineage of large filter-feeding species, the pachycormids, who reached maximum lengths of 52 feet. From the cartilaginous fishes emerged the first rays, which flattened their bodies and became bottom-trawling predators of hard-shelled invertebrates. Marine reptiles complemented their distant relatives, with the ichthyosaurs and plesiosaurs reaching greater importance. One lineage of plesiosaurs, called pliosaurs, became the major oceanic predators of the Jurassic, sprouting enormous heads lined with sharp teeth for ripping up fishes and other marine reptiles. They too, reached impressive sizes, with Pliosaurus funkei estimated at 27 feet in length.
In the air, the pterosaurs had continued their presence and dominion, and there were new varieties that exploited different food sources. Some fancied the seashores and had teeth like toothpicks for spearing small fish. Others became very small and chased after flying insects, snapping them in their bear-trap-shaped jaws. Still others seem to demonstrate changes brought about by sexual selection. In evolution, the concept of sexual selection states that organisms will compete with each other for the favor of their mates, who select the individuals that will be beneficial partners and good parents. Thus, it is different from natural selection, where it is the environment that selects for advantageous adaptations. Sexual selection has been able to explain the strange gallery of horns and antlers, long and colorful feathers, sails and spines, and other seemingly extravagant structures often found on living things. The large antlers of moose and elk, the colors and feathers of peafowl and pheasants, and the manes of lions were the result of this form of evolution. These features are sexually dimorphic, meaning that they are different between males and females and can be used to distinguish the two. The northern cardinal, for example, has the males a bright shade of red, while the females are a light brown. In pterosaurs, there is possible evidence that sexual selection was at work on some species, where the presence of head-crests seems to show a marked difference in size between sexes. It is likely that these structures (which would have certainly been brightly colored and patterned) were viewed by pterosaurs as indicators of overall health and physicality, and over time would have encouraged the production of even more extravagant crests.
On land, the dinosaurs had a strong hold as the major herbivores and carnivores on all terrestrial ecosystems. By now they had produced enormous forms and had begun to really diversify into all of their major lineages, including all the forms familiar with laypeople today.
A number of theropod lineages had evolved truly massive predators, seemingly in response to the larger prey species that were now available as a food source. These dinosaurs had adapted their front teeth to have more of a pointed shape than a simple vertical shaft, however, their skulls were often not strong enough to deliver hard, grabbing bites towards their prey. They were keener to run up to prey animals and slash at them with their open mouths, leaving deep cuts and selecting small bites of flesh, leaving the other dinosaurs to eventually succumb to blood-loss and death. These Jurassic theropods often had ornamental bumps and crests atop their heads too, perhaps another instance of sexual selection at work. Two familiar Jurassic theropods are the horned Ceratosaurus which sported a row of hard, bony bumps along its back, and the larger Allosaurus that had a wider chest and deeper face. By comparing these two theropods, you can tell some curious things about their behaviors. Ceratosaurus has particularly small arms with stubby fingers and short claws; meanwhile it’s jaws are adorned with very long teeth. Allosaurus’s arms were rather long, tipped with lengthy and hooked claws, while the teeth were shorter in its jaws. This implies that Ceratosaurus relied more on its mouth for taking down prey, while Allosaurus may have used its arms to latch onto prey animals while it slashed at them with its teeth. Incidentally, stress-tests on Allosaurus skulls show remarkable strength and its overall size suggests that it was going after larger animals than Ceratosaurus, which may have been content with medium-sized prey. These two animals did share the same environment and similar prey and would likely have avoided direct competition by hunting at different localities, similar to how lions and leopards interact today.
There was another group of theropods active during the Jurassic, one that is of great importance to many paleontologists. These are the coelurosaurs, and for much of their early history they remained small-bodied and certainly steered clear of the larger theropods in their ecosystem. Coelurosaurs are distinguished by an increase in brain size and a lengthening of the digits. There was a particular trait that perhaps evolved among the common ancestor of the dinosaurs that became very pronounced among the coelurosaurs: filamentous covering. These fur-like protofeathers would have acted as insulating structures that aided in keeping their internal body temperature warm and later on may have served as a template for display structures and colors (another outcome of sexual selection). One lineage of coelurosaurs, the tyrannosaurs, evolved towards the middle of the Jurassic Period. They had fused their nasal bones together to give them a much more powerful bite, but they were (for a time) small to medium-sized animals. Another lineage, the maniraptors, modified their arms to make them longer, their wrist became more specialized for folding against the body, and they changed their legs to allow a more crouched posture. Their protofeathers had now developed into proper feathers, with pennaceous (or quill-like) plumage that would have formed large wings on their forelimbs and rows along their tails. It was 165 million years ago that the direct ancestors of birds evolved from among the maniraptors. The most familiar of these animals is Archaeopteryx, but in all honesty they would have not looked very different from the other feathered coelurosaurs that shared the world with them. These were not flying animals at first, more content for short gliding and a mostly ground-based existence, though it appears possible that these early birds could flap their wings enough to remain balanced on prey items while they dispatched them or flapped their wings to run up vertical surfaces like tree-trunks; these are two behaviors that birds today do. Birds are, quite literally, avian reptiles.
The other, herbivorous dinosaurs were diversifying as well. By the end of the early Jurassic, the sauropods were all quadrupedal animals that supported their weight on columnar limbs tipped with padded digits. The hands gradually lost all but one claw, while the feet were supported by padded soles. The sauropods of the Jurassic grew to immense sizes and increased the lengths of their necks for higher browsing capabilities. Now the tree tops were no longer safe from plant-predators thanks to their height. Their nasal bones became enlarged, meaning that in life these dinosaurs would have sported massive nasal chambers. It’s unclear why this trait formed: paleontologists have tied them to better smelling, to the production of better calls, or to both. By the end of the Jurassic, two major groups of sauropods had evolved, distinguished by their general anatomy. The diplodocoids, of which the 70-foot Brontosaurus and the almost 100-foot Diplodocus belong, have long snouts with peg-shaped teeth which are all collected at the front of their jaws. This implies that these dinosaurs were using their mouths like rakes, scrapping up conifer needles and ferns. The macronarians, including the over 60-foot Brachiosaurus, are so named due to their boxy and blunt skulls with particularly large nasal openings. Their teeth ranged throughout their jaws (like earlier sauropods) and this meant that they engulfed more plant matter. In shape, diplodocoids were built slenderly and had long whip-like tails that may very well have been used like whips for defense, though the jury it out on that. Macronarians, in contrast, were bulky and built upwards, meaning that their long necks were always reaching upwards, like giraffes today.
Ornithischian fossils become more common during the Jurassic, and there is evidence that (like coelurosaurs) all their descendant groups had evolved from small, bipedal dinosaurs covered in filamentous fuzz. Though probably omnivorous at first, later groups shifted to herbivory. The ornithopods were among the first and only dinosaurs to develop proper chewing, whereby they ground up their food through the sliding of their teeth against each other, versus the other dinosaurs which simply bit up their food and swallowed it whole. Chewing allows for more nutrients to be gathered from plant food, so ornithopods became master herbivores and soon spread out across the globe. The group of ornithischians called marginocephalians is distinguished by their name-sake feature: a stretching and increasing in size of the bones behind the skull, which formed a great margin. They remained very small animals in their early evolution, using their sharp beaks to nip at plants and perhaps small animals as well. The third lineage of ornithischians that really owned the Jurassic were the thyreophorans or armored dinosaurs. They developed into primarily quadrupedal animals, with massive guts, thick legs, and, yes, armor plating on their bodies. One subset of this group was the stegosaurs, whose armor was transformed into a flattened and paired row of plates along their backs. These plates would have been useless as defensive organs, and so may have performed a role in sexual display. Their tails were lined with their own row of bony spines which, when swung, could deliver a deadly blow to any would-be predator. The other subset were the ankylosaurs who kept their body armor and took it to extreme levels, covering their faces in strong bone. These animals were low-browsers, mostly going after ferns and conifer brushes and lapping up the leaves with a massive, prehensile tongue.
The other reptiles of the Jurassic remained minor characters, but they were undergoing their own experiments. One lineage of the surviving pseudosuchians, the crocodylomorphs, started out as small, upright animals that ran through the underbrush with long, spindly legs. Later descendants included forms that entered the oceans and became marine reptiles, complete with finned tails and paddle-like limbs, and others that hung out near rivers and lakes as ambush predators. From the lepidosaurs finally emerged the tuataras (today confined to a single species in New Zealand) and the lizards, which included the first skinks, geckos, and snakes. Snakes, indeed, are legless lizards today, but the ancestral snakes of 170 million years ago still had reduced and insignificant limbs. Over time they would lose their forelimbs first and later their hindlimbs, but just why and where they lost their limbs is unclear. Paleontologists are torn between snakes losing their legs due to a burrowing lifestyle and snakes losing their legs because they evolved as aquatic animals. Meanwhile, among amphibians, frogs finally developed the ability to hop and truly lost their tails. The two other living groups of amphibians, the lizard-like salamanders and the legless, burrowing caecilians, are represented by very good fossils at this time, though they almost certainly evolved earlier in the Triassic.
Land arthropods experienced new evolution too, with spiders in particular undergoing a radiation of new species thanks to the evolution of spinnerets. There were new groups of insects appearing too: the first earwigs and the first moths, who had thin wings and only later developed their thin proboscis for sipping up water. The sawflies had given rise to the first wasps, which modified their plant-cutting ovipositor into a piercing organ or stinger. This tells us that wasps became predators that injected venom into their prey to subdue it. At the other end of the insect world, the earliest fleas included some dramatically large species up to an inch in length. These would not have been the jumping species we know today but rather lumbering, louse-like insects that would have taken bites out of mammals or small dinosaurs.
Jurassic mammals began to really explore new niches under the forests of gymnosperms and ferns that dominated the landscape. While some mammals remained small, shrew-like creatures like their ancestors, others became semi-aquatic (like beavers) or took to gliding (like flying squirrels) or went underground as burrowers (like gophers). What must be understood is that the main types of mammals today were not yet established during the Jurassic and that all these new mammals belong to now extinct lineages.
Part 3
The Jurassic ended with not so much as a bang but a whimper, as a few land and sea faunas faced some displacement as new lineages rose and took over. The final period of the Mesozoic is the Cretaceous Period, 145 to 66 million years ago. The continents continued to break apart and move in opposite directions, with Gondwana’s rifting pretty much complete by 110 million years ago. The Atlantic Ocean now stretched from the tip of North America’s eastern seaboard to the bottom of present-day Tierra del Fuego. Incidentally, with the movement of South America away from Africa and its subsequent contact with the Pacific plate, the Andes mountains began to form. Around 96 million years ago, Madagascar separated from India, and not long after Sahul split from Antarctica. By the end of the Cretaceous, the continents had mostly been near to their present locations, but the general geography of the globe was still vastly different. For one, sea levels rose significantly, with their average height at 557 feet. This meant that large parts of the Earth’s surface were now covered by shallow seas, with North America, Amazonia, Siberia, and Saharan Africa in particular sporting wide interior seaways by 100 million years ago. Europe was still an archipelago, but so was Antarctica.
With so much water to go around, marine ecosystems really flourished. In a continuation of events that began during the Jurassic, the Mesozoic Marine Revolution was reshaping the basic composition of the ocean’s ecology. Red algal phytoplankton remained the dominant marine photosynthesizing organisms, with the shelled diatoms in particular undergoing a spike in diversity. The coccolithophores were doing well too, and it was now that great quantities of their deceased began to form serious chalk deposits around the world. The beautiful and iconic White Cliffs of Dover in southern England, are formed of Cretaceous coccolithophores. Among marine invertebrates, the crinoids and brachiopods that so characterized Paleozoic faunas had by now truly been relegated to their present-day positions as deep-water curiosities. The main stars of the seafloor were the bivalve and gastropod mollusks, the sea stars and urchins, and the crabs and lobsters. The most prominent feature of these new aquatic animals was their adaptations towards the consumption of hard-shells, with many species developing the ability to pierce or crush through shells, which in turn caused these prey animals to strengthen their hard-coverings or bring about new defenses, like poison stingers or a shift in environment towards more deep-sediment burrowing. Many iconic marine animals, like the cone snail, the conch, the clawed lobster, the hermit crab, and the heart urchin evolved during the Cretaceous. Of importance was another, temporary shift in reef-building organisms. While the scleractinian corals were still present, and themselves diversifying, it was a group of bivalve mollusks called rudists that would form the reefs of the Cretaceous. Rudists were enormous filter-feeders that attached themselves to surfaces and formed great swarms along the seafloor.
Patrolling the waters above these invertebrates were their vertebrate neighbors. Ray-finned fishes had produced a number of new lineages, including the first cod and anglerfish. By now, modern-type sharks had evolved, with the mackerel-shark group (which today includes makos, sand tigers, and the great white) gaining a reputation as efficient marine predators. But there was no competition that the dominant members of the ocean food-webs were the marine reptiles. Around 90 million years ago, the ichthyosaurs had died out due to changing environmental conditions, and the pliosaurs went extinct not long after. This left a group of seashore adapted lizards, of unclear relation to living forms, to take up the niche of apex marine predator. These animals, the mosasaurs, were remarkable in their own right, adapting their limbs and tails into paired flippers and fins and developing an additional row of teeth in their palate, flanked by their already sharp and intimidating fangs. While some mosasaurs were average-sized reptiles, comparable to the ichthyosaurs that came before, a few were giants of over 50 feet. They no doubt preyed upon the plesiosaurs, who themselves were going strong and produced great, long-necked species, as well as another new group of marine reptiles, the sea turtles. Sea turtles were content to feast upon aquatic plants, soft-bodied fishes, and sea jellies with sharp beaks, and one kind called Ocepechelon sucked up its food with a bizarre turkey baster-like snout. Of note is the fact that sea turtles reached some immense sizes during the Cretaceous, with the marine Archelon sporting a 15-foot shell.
There were more pterosaurs during the Cretaceous than at any other point in the Mesozoic, and with a much more widespread diversity of niches and body-types. There were the ornithocheiraens, with pencil-like teeth in jaws marked by parallel crests. The tapejarids, with toothless beaks but enormous crests that often overshadowed their skulls, who may have been arboreal, fruit-eating pterosaurs. There was Pterodaustro, with a row of frilly-teeth in its lower jaw that must have sieved microscopic food from the water like a flying whale, and Pteranodon, who was an ocean-going predator that dived after fish with its toothless beak. Most iconic of all (and perhaps most popular among paleontologists) were the azhdarchid pterosaurs. These were stork-like animals that often walked along the ground in a sort of quadrupedal creep, poking around for small to medium-sized vertebrate prey. This lineage begat the largest flying animals that ever lived: found in Romania on what would have been an island in Europe’s archipelago was Hatzegopteryx with an estimated wingspan of around 33 feet, comparable to a Cessna aircraft.
On land, other reptiles continued to have a stable existence. Snakes and other lizards moved through the underbrush, with some occasionally finding themselves near a dinosaur nest and snagging a baby or two. Turtles gave rise to a few new forms, including the soft-shelled turtles and the pond turtles, who survive to the present day. Crocodylomorphs were giving rise to some brand-new groups, including herbivorous forms with mammal-like teeth, forms with armored bodies like armadillos, and duck-billed forms. They also produced some giants, including the slender-snouted Sarcosuchus that spanned over 38 feet. The modern members of the group, crocodiles and alligators, didn’t show up until 83 million years ago, meaning that – like sharks – the claim that the entire group remained unchanged since their origin is inaccurate as well. Among the amphibians, the ancestors of modern frog, salamander, and caecilian lineages were developing alongside other bizarre experiments, including the giant carnivorous Beelzebufo that certainly deserved that name.
One of the last major events in botanical history underpinned the Cretaceous World. Though their relatives were present in the Permian Period, and early members of this group appear to have evolved during the middle of the Jurassic, they radiated in such numbers that they directly affected most of the ecosystems of the Cretaceous world. These were the flowering plants, or angiosperms. Flowers are essentially reproductive organs, and an extension of the system that gymnosperms developed: a flower is just a modified stem and the petals were originally the leaves on that stem. Once pollen comes in contact with the flower's stigma (the elongated-bushy part surrounded by petals), they reach the ovaries located inside the stem and fuse with it. This fusion eventually leads to the creation of the seed. The question of how this process came to be is riddled with controversy, but the answer seems closely tied with the history of insects.
During the Cretaceous, most of the remaining insect lineages around today had evolved, including the first praying mantises, which evolved from insects related to cockroaches. Descended from among the cockroaches themselves were the first termites, which can digest cellulose with the aid of symbiotic bacteria in their guts. Termites form complex societies inside wood and in large mounds of dirt where they build their nests. A large, wingless queen is the sole insect that produces offspring, which develop into any number of specific morphs, including sharp-jawed, defending soldiers and food-collecting and nest building workers. These insects, though vast in numbers, essentially function as one single organism: they are eusocial. The wasp lineage evolved a number of different eusocial groups as well, including the first bees, ants, and hornets. Bees, in particular, are notable because they’re pollinators of flowering plants today. Also given that the evolution of bees and the radiation of angiosperms occurred at the same time, it seems that the two lineages of organisms co-evolved together. They’re certainly co-dependent: bees and other pollinating insects usually are adapted to feasting on the nectar that certain flowers produce, and most flowering plants rely solely on certain pollinating insects to reproduce.
The earliest flowering plants were weedy, bushy things that grew around the shores of lakes and ponds, meaning that they were possibly aquatic plants like the false dragonhead or the parrotfeather of today. One kind, Archaefructus, sported small tufts of leaves above the reproductive organs (there’s no petals or stigma to be seen). It is likely that the earliest flowering plants encouraged flying insects to their developing petals via bright colors, which the insects saw with their ultraviolet eyesight. Overtime, angiosperms moved outwards towards other inland environments, including the undergrowth of gymnosperm forests, which were still the dominant plant species. By 80 million years ago, however, angiosperms flipped the script and outcompeted the gymnosperms in many regions. A whole host of familiar and common plants, including members of the rose, magnolia, oak, dogwood, palm, and hickory families. And among these plants were the first fleshy fruits.
Most people can tell when fruit has ripened for consumption, and that is one of the adaptations that allowed angiosperms to be so productive and widespread. The bright and attractive colors of fruits tell animals that their seeds are ready to be dispersed, and these seeds are tough enough to withstand digestion. Once the animal in question defecated, the seed was planted in its own fertilizer. But one question remained, what animals were flowering plants encouraging with their edible fruit?
The mammals at the time were still minor members of the Cretaceous landscape, but by the beginning of this period, the three living lineages had evolved: the monotremes, who retained their egg-laying abilities and are represented today by the platypus and echidnas; the marsupials, who are live-birthing mammals but shift the later growth of their embryos to a pouch that contains the nipples; and the placentals, also live-birthers, but this time retain the young inside their uterus until it is more developed. Though there were certainly seed- and berry-eaters among them, the sheer size of the other fruits meant that this food source would not have meant much to them. Incidentally, there was an herbivorous terrestrial crocodylomorph, Simosuchus, alongside the fruit-eating tapejarids, but these were not widespread enough to encourage this evolution. Therefore, paleontologists have turned to the one lineage of reptiles that was still going strong and just so happened to reach their zenith during the Cretaceous: the dinosaurs.
Part 4
The group that must have really encouraged the spread of soft fruit-bearing angiosperms was the ornithischians who, having evolved into their major lineages, rose to prominence during the Cretaceous. The stegosaurs held on until roughly 100 million years ago, but by the time they went extinct their relatives the ankylosaurs had given rise to many types. Some members of this group, like Edmontonia and Sauropelta, lacked the defensive tail-club that their kin had and instead developed their neck and body armor or osteoderms into projecting spines. This was clearly more of a defensive anatomy and any attacking predator risked a slash on their leg. Other ankylosaurs, like Euoplocephalus and Talarurus, had moved several of their osteoderms towards the tail-end of their… well… tail, and formed a reinforced bony-club. Like the tail-spines of the earlier stegosaurs, these tails could be swung at will towards predators, and biomechanical studies demonstrate that these animals could hit a target and generate an impact stress of between 364-718 megapascals. For larger ankylosaurs, this was enough to outright shatter bones, and the resulting infection of a wound could very well kill an attacking theropod. Porcupines rely on this strategy today, and many carnivorous mammals that have been dealt a spiny blow find themselves dead later on.
While the ankylosaurs rooted along the ground for low-growing plants, they would have been joined by the two lineages of marginocephalians, who had by the Cretaceous distinguished themselves. The pachycephalosaurs were bipedal dinosaurs that thickened the tops of their skulls with solid bone. The characteristic margin formed a shelf behind their heads that was often decorated with pointed bumps and horns. The purpose of this hard head has been speculated by many to function like the horns of sheep and goats, who butt heads during the mating season as a test of strength. This would mean that these domed structures were the result of sexual selection. Interestingly, some paleontologists have compared the skulls of different species of pachycephalosaurs and concluded that the young, born without the dome, underwent changes during sexual maturity by which the skull acquired the thickened bone (again, similar to the growth stages that sheep and goats undergo). This is still a debated subject.
The other group of marginocephalians were the mostly quadrupedal ceratopsians, who are familiar by the popular Triceratops. This was one of the last and largest members of the group to evolve, and just the tip of the iceberg of the total diversity of these dinosaurs. In ceratopsians, the beak was enlarged in size and formed a sharp downward curve, almost like that of a parrot or finch. The margin here was greatly elongated into a solid or hollow frill that would have extended and covered the neck and upper back, but due to the apparent fragility of this flattened board of bone, it’s unlikely that this was used for protection. The sheer size of the frill lends a possibility that this was an efficient sexual display structure that may have been brightly colored and or patterned. The real defense was found on the horns that grew atop the animal’s heads. These horns came in a variety of styles, with some species sprouting two large brow horns and a smaller nasal horn, like Triceratops as well as Chasmosaurus and Pentaceratops, and others developing a long nasal horn and smaller brow horns, like Centrosaurus and Styracosaurus, the latter displaying a crown of thorns along its frill. It might seem that these structures were efficient at goring the stomachs of attacking predatory dinosaurs, and though they certainly would have been, their main function appears to have been as display structures. So, yes, sexual selection would have been at work again. Of the studies we have on ceratopsians, it appears that the horns began as small bumps on the heads of young dinosaurs, only later lengthening in size as they grew. If one examines living mammals, like deer, cattle, and antelope, their horns and antlers always begin as diminished structures, only later reaching full size as they mature and begin fighting for mates, so it would not be a stretch to argue that the same thing was happening among ceratopsians. So, a Triceratops would more likely face off against another rival Triceratops than a Tyrannosaurus.
Ornithopods, like other ornithischians, reached their greatest diversity in the Cretaceous and among them diverged a lineage with elongated and slightly flattened snouts, longer hands, and an extension of the chewing teeth into a more complex, grinding battery. These were the hadrosaurs, often known as duck-billed dinosaurs, though given that in life their snouts would have been covered by a blunt, chopping beak, they wouldn’t have really looked duck-like in their face. Nor were they duck-like in behavior: all evidence available tells us that hadrosaurs were terrestrial animals that formed large herds that swept through the lands of the Cretaceous, grazing on ferns and other low- to medium-growing plants. They grew to massive sizes too, with the largest members of the group surpassing 50 feet in length, and given the lack of horns or claws that would have been enough to deter most predators. In what could be another instance of sexual selection at play, many members of this group developed remarkable crests atop their heads, similar to what some pterosaurs had done. Unlike the flying reptiles, hadrosaur crests were hollow and housed an extension of their nasal passages. Biomechanical studies suggest that these dinosaurs could produce loud calls with their crests, similar to the noises of wood-wind instruments, and these no doubt were complicated by whatever additional soft-tissues were present. Given the wide variety of crest shapes, they would have definitely been brightly colored and used to attract mates.
The sauropods experienced their own radiation of forms, and around 125 million years ago they gave rise to the titanosaurs. These were, without a doubt, the largest land animals that ever walked the Earth. In the last few years a number of finds have revealed enormous leg bones or vertebrae and, comparing them to more complete remains, these have allowed paleontologists to estimate their size. These dinosaurs, with powerful names like Dreadnoughtus and Patagotitan, may have been up to 121 long and weighted as much as 76 metric tons. While talk of their size is certainly invigorating, what truly made titanosaurs and other sauropods big was their ecological impact. Like elephants today, herds of sauropods would have pushed through the conifer forests of the Cretaceous, eating as much of the needle-like leaves as they could and then leaving nothing but broken and trampled trees behind. This would have kept the forests from overgrowing and subsequently encouraged new growth of other plants, as well as sapling trees. In turn, this allowed other, smaller dinosaurs to find the food that they needed, and their predators would follow suit. While they were certainly common animals, titanosaur numbers were controlled through their breeding and through their parenting strategy, or rather, lack of parenting. Titanosaurs, and other sauropods, appear to have been R-selected species: laying enormous quantities of eggs and subsequently leaving them to fend for themselves, with the understanding that at least some would survive to adulthood. Sea turtles are living analogs, with their young undergoing an intense and harsh childhood with a barely-present chance that all will live to see another day. Paleontologists have recognized that titanosaurs would have been R-selected species because we have discovered vast nesting grounds that belonged to titanosaurs: there is no way to justify that all these eggs and offspring were cared for individually. For comparison, K-selected species physically raise a small number of offspring and ensure that all or most of them have the skills necessary for survival in the wild. For mammals like us, we are K-selected organisms.
The last major group of dinosaurs, the theropods, had expanded into a number of new niches. Many groups of theropods developed into omnivores or herbivores, and either lost their teeth or shortened and blunted them for cropping plants. Some of these, the ornithomimosaurs, evolved into ostrich-like animals and definitely resembled ostriches with long tails and working fingers. Their lengthy necks would have been good for reaching medium-growing plants and their legs gave them an impressive stride, meaning that they must have been ostrich-like in their speed too. A smaller group, called the alvarezsaurs, were much smaller in stature. They had all but lost most of their fingers – save for their thumbs – and greatly reduced the length of their arms. What on earth these dinosaurs were doing is unclear, but an insectivorous lifestyle has been suggested. Just as bizarre were the therizinosaurs, which extended the length of their arms and claws and developed a cropping beak and back teeth. These were certainly herbivores, using their sharp claws to bring down the branches of trees and squatting down on their haunches as they ate like some sort of avian panda bear.
There were plenty of carnivorous theropods as well, like the tyrannosaurs. By the later end of the Cretaceous, they had all developed into larger and more powerful predators. Their heads increased in size, with their palate hardening, their teeth thickening, and the roots of those teeth extending deeper into their sockets. While this was going on, their arms had shrunken, and they lost a finger too, leaving only two digits on their hands. This implies that they were relying more on the head for dispatching prey than the forelimbs. Not to mention that their hindlimbs were toughened by a modification of the middle-most foot bone that had it ‘pinched’ between its flanking bones. This arctometatarsus allowed tyrannosaurs to move faster and more efficiently. Some members of this group, like Albertosaurus and Alioramus, would have been fleet-footed killers, able to run down their prey, but the posterchild of the group, Tyrannosaurus, was a heavy-bodied predator that has been estimated to have only cruised as fast as 12 miles per hour as adults. Spanning 40 feet in length and weighing at least 8 tons, Tyrannosaurus would have been able to tackle armored animals, crushing through their bones with as much as 57,000 newtons of force. One tantalizing fossil demonstrates that an individual managed to bite through the horn of a Triceratops, direct evidence that it sometimes had to face prey head on.
At the other end of the theropod spectrum were the dromaeosaurs. These are the popular ‘raptors’ from the Jurassic Park films, though their film presence as pack-hunting, hyper killers undermines what their actual behaviors would have been. Biomechanical studies and comparative research with living species tell us that dromaeosaurs behaved very similarly to modern birds-of-prey (incidentally, known as raptors). These dinosaurs would have stalked smaller animals, launching themselves atop their bodies, grabbing onto their flesh with their raised and curved toes, stabilizing themselves with their winged arms, and ripping into them with their sharp-toothed mouths. Indeed, if you found yourself in the Cretaceous, you would be alive when they started to eat you. Related to the dromaeosaurs were the birds, who following the Jurassic had diversified into a number of new forms. Cretaceous birds shortened their bony tails in favor of a pygostyle, where the remaining vertebrae fused together. A few managed to lose their teeth in favor of a beak, but the majority still retained them. Some still had working fingers with claws on their hands, while others sealed them together into one structure. One lineage of birds, the Hesperornithes, became marine animals, nearly losing their forelimbs as they adapted their hindlimbs into paddling organs with webbed toes. They were essentially penguins before penguins were a thing.
The Cretaceous was a blossoming time for living things, with the seas full of ammonoids, fishes, and marine reptiles, the air soaring with pterosaurs and early birds, and the land brimming with flowering plants, scurrying mammals, and a host of ruling reptiles. But things took a serious turn of events and, mirroring the Paleozoic before it, the Mesozoic ended with a mass extinction event 66 million years ago. This is perhaps the most familiar of these great losses of life, but what may surprise many are the intricate details that underpinned this event, and the amount of evidence we have for how things really went.
It has become clearer in recent years that the final million years of the Cretaceous Period was a time of strong changes on Earth. The high sea levels that characterized the age had lowered, and the great interior seaways that swept through Africa, Asia, and the Americas receded. The archipelago that was Europe was no more, now those lands were connected in a proper subcontinent. Given that many parts of the land were reconnected, animals were able to move between them, not to mention that now the faunas and floras that characterized these inland seas had lost their habitats and were now pushed outwards into the open ocean. This was an event known as the Maastrichtian Regression, named after an age in the Late Cretaceous Epoch: the last age of the Mesozoic. Groups of organisms at some localities appear to be at a lower diversity than previously found, though it is controversial whether this was the case everywhere, or even whether the drop in sea levels was to blame.
On the Indian subcontinent, which was during the Cretaceous its own island, there began a series of periodic volcanic eruptions right at the end of the period that brought vast quantities of lava to the surface. The rocks this eventually formed, called the Deccan Traps, suggest that the coverage was on an unimaginable magnitude, with over 470,000 miles of lava being pooled across India. As grand, miles-long volcanic eruptions tend to do, the carbon output would have been pretty severe, and that’s not to forget the periods of darkness that would have followed as ash clouds blotted the sun in places (bringing periods of intense cool to punctuate the hot times) and the instances of acid rain that poisoned the air and water. Clearly, this really was a difficult time for living things but there was an additional insult to injury coming out of the vastness of space.
During the 1980s and onwards, geologists noticed a thin band of iridium occurring at many sites around the world, always bordering the end of the Mesozoic Era. As far as minerals go, iridium is not common on Earth, but prime sources can be found in extraterrestrial rocks, like meteors. This led to the proposal that the planet had been impacted by a bolide at the end of the Cretaceous, 66 million years ago. One question remaining was, where was the crater? It took researchers many years until 1990, when a proper connection was placed between the iridium band and a 110 mile wide crater in the Yucatan Peninsula that, though sealed with Earth, left traces of its impact in the remains of shocked quartz and other minerals that indicate a severe crash took place. The rocks of the crater were properly dated, and the case was sealed: the Cretaceous Period ended with a bolide impact. How this event unfolded would have made even the most hard-core disaster films look tame.
There would have been no warning, maybe the presence of a new star in the sky that later turned into a second Sun. But on the day that it struck, it hit the Earth with enough force to push the atmosphere away and send miles of rock into outer space. Imagine setting off 100 trillion tons of TNT. The sheer size of the impact, which would have been in the ocean, sent off enormous tsunamis with waves many hundreds of feet high. Earthquakes would have radiated from the site and certainly any organisms within the range of the blast would have just vanished. The long-term effects of the bolide impact would have caused a particularly intense period of global cooling, like a nuclear winter, as a huge eternal fog of dust blanketed the planet and destroyed all ecosystems. An already impacted biosphere was just now dealt a harsh blow, with 75% of all organisms going extinct. Of course, all of the dinosaurs save for one lineage of birds were killed off, but the great host of marine reptiles – the plesiosaurs, mosasaurs, and giant sea turtles – and the flying pterosaurs were wiped out too. All the rudist bivalves that formed great reefs and the great clusters of coiled ammonoids and squid-like belemnoids were all gone, and there were losses among the mammals, reptiles, insects, and gymnosperms too. It was another tragic end to a time of amazing and beautiful organisms, but as is the case for the story of life on Earth, it was time for another era to take hold and share its own amazing and beautiful plants and animals.
And with that, we must lay anchor to our river journey. In the next episode, we close out our story of life with the Cenozoic Era and visit a world where the survivors of the Cretaceous extinction event, including our mammalian relatives, filled the planet with their descendants. We see the rise of many popular and well-known animals to our societies and see how many of the landmarks we know-and-love came to be. Grasslands form and expand, mountains rise and fall, and the stage sets for the evolution of our ancestors.
That’s the end of this episode of On the River of History. I’d like to thank my friend and PhD student in paleontology Albert Chen for reading over this transcript and checking it for accuracy. You can find him on Twitter @albertonykus and you can check out his blog Raptormanics. If you enjoyed listening in and are interested in hearing more, you can visit my new website at www.podcasts.com, just search for ‘On the River of History’. This podcast is also available on iTunes, just search for it by name. A transcript of today’s episode is available for the hearing-impaired or for those who just want to read along: the link is in the description. And, if you like what I do, you’re welcome to stop by my Twitter @KilldeerCheer. You can also support this podcast by becoming a patron, at www.patreon.com/JTurmelle: any and all donations are greatly appreciated and will help continue this podcast. Thank you all for listening and never forget: the story of the world is your story too.
1 note
·
View note
Link


Un articolo pubblicato sulla rivista "Scientific Reports" riporta la scoperta di due nuovi esemplari di Eretmorhipis carrolldongi, un rettile marino che visse circa 250 milioni di anni fa, nel periodo Triassico Inferiore, nell'odierna Cina. Un team di ricercatori ha studiato questi nuovi esemplari con particolare interesse perché dei precedenti uno aveva il cranio ma era in un cattivo stato di conservazione e l'altro era privo di cranio. I nuovi esemplari mostrano che questo rettile aveva similitudini con i moderni ornitorinchi, con una sorta di becco cartilagineo e recettori che gli permettevano di cacciare basandosi sul tatto.
0 notes
Last Seen Blogs

glitterybeardonut
Gino Concept Store

roahnai
RoahNai

understeerking
UndersteerKing

gerloveless
Ash

ladypalta
aza es mi droga